
Está usted aquí: inicio > genes ndh
(Puede consultar el documento en formato pdf haciendo clic en este enlace)
EVOLUCIÓN Y EXPRESIÓN DE LOS GENES ndh
El DNA de cloroplastos de la mayoría de las plantas incluye 11 genes ndh que codifican polipéptidos del complejo Ndh, el cual está implicado en la regulación del nivel redox de los transportadores fotosintéticos de electrones para optimizar el flujo cíclico y la fotofosforilación. La síntesis de los polipéptidos NDH (codificados por los genes ndh) y los niveles del complejo Ndh aumentan especialmente en situaciones estrés fotooxidativo y senescencia.
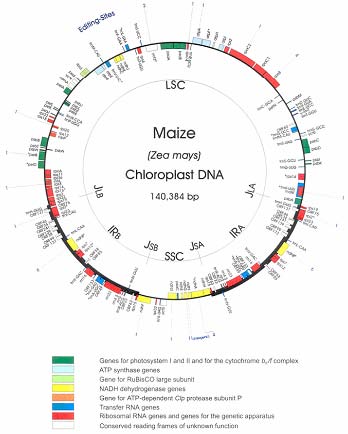
Como muestra el mapa del genoma de maíz en amarillo, seis genes ndh
(H, A, I, G, E y D) se agrupan en el
operón ndhH-D que también incluye el gen psaC
(entre ndhE y ndhD) para el polipéptido PsaC del fotosistema I.
Tres genes (ndhC, ndhK y ndhJ)
forman un segundo operón, mientras que ndhB (duplicado) y ndhF forman
probablemente sendas unidades monocistrónicas.
Los genes ndh no se encuentran en el DNA de cloroplastos de la mayoría de las algas: Glaucofitas
(Cyanophora paradoxa), Criptofitas
(Guillardiatheta), rodófitas
(Cyanidioschyzon merolae, Cyanidium caldarium,
Porphyra purpuea), Euglenofitas
(Euglena gracilis, Euglena longa), cromofitas (Odontella sinensis) y algunas
Clorofitas
(Chlamydomonas reinhardii, Chlorella vulgaris).
Pero si se encuentran en el DNA de cloroplastos de algunas Clorofitas
(Chaetosphaeridium globosum,
Nephroselmis olivacea,
Mesostigma viride) y casi todas las Briofitas, Pterdofitas, Gimnospermas y Angiospermas.
Excepciones entre plantas superiores incluyen algunas fotosintéticas
(Pinus koraiensis,
Pinus thunbergii) y algunas no fotosintéticas (
Cuscuta reflexa,
Orobanche minor) que normalmente tienen pseudogenes ndh.
Transcritos del operón ndhH-D
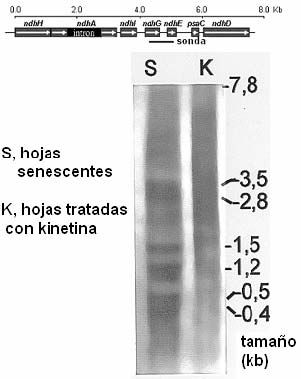

Los transcritos de los genes ndhB y ndhF aumentan durante el envejecimiento y en respuesta al estrés fotooxidativo. Los operones policistrónicos con genes ndh presentan un perfil complejo de transcritos cuya abundancia relativa varía, en general aumentando las formas monocistrónicas durante la senescencia foliar. En el caso más estudiado, del operón ndhH-D, el análisis de transcritos revela múltiples etapas de procesamiento desde un transcrito primario de 7,8 kb y que incluyen ediciones de algunas bases C a U, eliminación del intrón del ndhA y cortes aproximadamente intergénicos.
La edición de los transcritos de los genes ndh consiste, como para los transcritos de otros genes plastidiales de plantas superiores, en la conversión de algunas C en U. Con mucha diferencia, los genes ndh concentran la mayor parte de sitios editados de los genes plastidiales de angiospermas. Posiblemente, en la línea que dio origen a angiospermas los genes ndh eran prescindibles y acumuló diversas mutaciones pero no llegó a eliminarlos. La bajada de la concentración de CO2 y la invasión de nuevos espacios que hacían más frecuentes situaciones de estrés fotooxidativo y, posiblemente también, la emergencia de hábitos caducifolios asociados con senescencia foliar propiciaron el rescate funcional de los genes ndh. En aquellos cuyas únicas mutaciones consistieron en transiciones genómicas T a C, tal rescate fue posible por edición a nivel de transcritos de esas C a U que permitían la codificación de aminoácidos críticos, muy conservados cuando se comparan alineamientos de las secuencias de las proteínas correspondientes. Posteriormente, en la evolución de angiospermas, sitios de edición van desapareciendo por retro-mutación C a T a nivel genómico.
Se conoce poco de la maquinaría de edición y del orden y control del procesamiento de transcritos policistrónicos de cloroplastos. En el caso del operón ndhH-D casi todos los sitios son editados en las primeras etapas del procesamiento. Un excepción importante es el sitio III del gen ndhA que solo es editado después de perder el intrón. En realidad, una población significativa de transcritos contiene el intrón del ndhA con todos los sitios ya editados excepto el citado sitio III (en el segundo exón y próximo al intrón). Predicciones estructurales indican que la típica estructura secundaria necesaria para la eliminación del intrón tipo II mantiene en un brazo apareada la C del sitio III con una G. Solo cuando se ha eliminado el intrón queda libre la C y es editada a U. Cabe pensar que la necesidad de esta C para el procesamiento del intrón, ha impedido la restauración genómica de la misma por T y, en consecuencia, la persistencia como tal sitio III que requiere edición en muchas plantas.
Otro hecho singular del procesamiento de transcritos del operón ndhH-D es el mecanismo por el que se acumulan altos niveles del transcrito monocistrónico del gen psaC en contraste con los bajos niveles de transcritos de genes ndh. Los cloroplastos contienen entre 100 y 200 veces más polipéptido PsaC (componente del fotosistema I) que proteínas NDH. En buena parte, esto se debe a que durante el procesamiento del transcrito primario del operón ndhH-D se acumula un transcrito mocistrónico del psaC estabilizado por prolongar su 3´-terminal abarcando secuencias del gen siguiente ndhD lo que, de paso, determina una menor abundancia del transcrito de este último. Es así evidente que un mejor conocimiento del procesamiento de los transcritos del operón
ndhH-D aportará importantes claves sobre el control de la expresión de los genes ndh y, en general, de los mecanismos de control post-transcripcional en cloroplastos.
